Innhold
- Oppdagelsen av DNA
- DNA og arvelige trekk
- Strukturen til DNA
- Nukleotider og nitrogenbaser
- DNA-replikasjon
DNA er en av de få kombinasjonene av bokstaver i kjernen av en vitenskapelig disiplin som ser ut til å vekke et betydelig forståelsesnivå, selv hos personer med liten levetidseksponering for biologi eller vitenskapene generelt. De fleste voksne som hører uttrykket "Its in her DNA", gjenkjenner umiddelbart at en bestemt egenskap er uatskillelig fra personen som blir beskrevet; at karakteristikken på en eller annen måte er medfødt, aldri forsvinner og er i stand til å bli overført til den personenes barn og videre. Dette ser ut til å stemme selv i hodet til de som ikke aner hva "DNA" til og med står for, som er "deoksyribonukleinsyre."
Mennesker er forståelig nok fascinert av konseptet med å arve trekk fra foreldrene og overføre sine egne egenskaper til deres avkom. Det er bare naturlig for folk å gruble på sin egen biokjemiske arv, selv om få kan forestille seg det i så formelle vendinger. Erkjennelsen av at ørsmå usettede faktorer i hver av oss styrer hvordan folk ser ut og til og med oppfører seg, har sikkert vært til stede i mange hundre år. Men først på midten av 1900-tallet avslørte moderne vitenskap i strålende detaljer ikke bare hva molekylene som var ansvarlige for arv, men også hvordan de så ut.
Deoxyribonucleic acid er faktisk den genetiske blå alt levende ting opprettholder i cellene sine, en unik mikroskopisk finger som ikke bare gjør hvert menneske til en bokstavelig en-av-en-type individ (identiske tvillinger unntatt for nåværende formål), men avslører mye viktig informasjon om hver person, fra sannsynligheten for å være relatert til en annen spesifikk person til sjansene for å utvikle en gitt sykdom senere i livet eller overføre en slik sykdom til kommende generasjoner. DNA har ikke bare blitt det naturlige sentrale punktet i molekylærbiologi og i livsvitenskapen som helhet, men også en integrert del av rettsmedisinske vitenskap og biologisk prosjektering.
Oppdagelsen av DNA
James Watson og Francis Crick (og mindre ofte Rosalind Franklin og Maurice Wilkins) får bred kreditering av oppdagelsen av DNA i 1953. Denne oppfatningen er imidlertid feil. Kritisk sett konstaterte disse forskerne at DNA eksisterer i tredimensjonal form i form av en dobbel helix, som egentlig er en stige som er vridd i forskjellige retninger i begge ender for å skape en spiralform. Men disse målbevisste og ofte feirede forskere bygde "bare" på det møysommelige arbeidet til biologer som slet på jakt etter den samme generelle informasjonen helt tilbake til 1860-årene, eksperimenter som var like banebrytende i seg selv som Watson, Crick og andre i tiden etter 2. verdenskrig.
I 1869, 100 år før mennesker skulle reise til månen, prøvde en sveitsisk kjemiker ved navn Friedrich Miescher å trekke ut proteinkomponentene fra leukocytter (hvite blodlegemer) for å bestemme sammensetning og funksjon. Det han i stedet hentet ut kalte han "nuclein", og selv om han manglet instrumenter som trengs for å lære hva fremtidige biokjemikere ville kunne lære, skjønte han raskt at denne "nuclein" var relatert til proteiner, men ikke selv var protein, at den inneholdt en uvanlig mengde fosfor, og at dette stoffet var motstandsdyktig mot å bli degradert av de samme kjemiske og fysiske faktorene som degraderte proteiner.
Det skulle gå over 50 år før den virkelige betydningen av Mieschers-arbeid først ble tydelig. I det andre tiåret av 1900-tallet var en russisk biokjemiker, Phoebus Levene, den første som foreslo at det vi i dag kaller nukleotider, besto av en sukkerdel, en fosfatdel og en basedel; at sukkeret var ribose; og at forskjellene mellom nukleotider skyldtes forskjellene mellom basene deres. Hans "polynukleotid" -modell hadde noen mangler, men etter dagens standard var den bemerkelsesverdig i mål.
I 1944 var Oswald Avery og kollegene ved Rockefeller University de første kjente forskerne som formelt antydet at DNA besto av arvelige enheter, eller gener. Etter å ha fulgt opp arbeidet så vel som for Levene, gjorde den østerrikske forskeren Erwin Chargaff to viktige funn: en, at sekvensen av nukleotider i DNA varierer mellom organismer, i motsetning til hva Levene hadde foreslått; og to, at i hvilken som helst organisme var den totale mengden nitrogenholdige baser adenin (A) og guanin (G) kombinert, uansett art, praktisk talt alltid den samme som den totale mengden cytosin (C) og timin (T). Dette førte ikke helt til at Chargaff konkluderte med at A-par med T- og C-par med G i alt DNA, men det hjalp senere til å styrke den konklusjonen som andre hadde nådd.
Til slutt, i 1953, samlet Watson og hans kolleger, som dra nytte av raskt forbedrede måter å visualisere tredimensjonale kjemiske strukturer, alle disse funnene sammen og brukte pappmodeller for å slå fast at en dobbel helix passet alt som var kjent om DNA på en måte ingenting annet kunne.
DNA og arvelige trekk
DNA ble identifisert som det arvelige materialet i livings-ting i god tid før strukturen ble avklart, og som ofte i eksperimentell vitenskap, var dette viktige funnet faktisk tilfeldig for forskernes hovedformål.
Før antibiotikabehandling dukket opp på slutten av 1930-tallet, krevde smittsomme sykdommer langt mer menneskeliv enn de gjør i dag, og å avdekke mysteriene til de ansvarlige organismene var et kritisk mål i mikrobiologisk forskning. I 1913 begynte den nevnte Oswald Avery arbeidet som til slutt avslørte et høyt innhold av polysakkarid (sukker) i kapsler av pneumokokk-bakteriearter, som hadde blitt isolert fra lungebetennelsespasienter. Avery teoretiserte at disse stimulerte antistoffproduksjon hos smittede mennesker. I England utførte William Griffiths i mellomtiden arbeid som viste at døde komponenter av en slags sykdomsfremkallende pneumokokk kunne blandes med de levende komponentene i en ufarlig pneumokokk og produsere en sykdomsfremkallende form av den tidligere ufarlige typen; dette beviste at uansett hva som beveget seg fra de døde til de levende bakteriene, var arvelig.
Da Avery fikk vite om Griffiths resultater, satte han i gang med å utføre renseforsøk i et forsøk på å isolere det presise materialet i pneumokokkene som var arvelige, og innlemmet i nukleinsyrer, eller nærmere bestemt nukleotider. DNA ble allerede sterkt mistenkt for å ha det som da populært ble kalt "transformerende prinsipper", så Avery og andre testet denne hypotesen ved å eksponere det arvelige materialet for en rekke stoffer. De som er kjent for å være ødeleggende for DNA-integriteten, men ufarlige for proteiner eller DNA, kalt DNAaser, var tilstrekkelige i høye mengder til å forhindre overføring av egenskaper fra en bakteriegenerasjon til den neste. I mellomtiden gjorde proteaser, som avdekker proteiner, ingen slike skader.
Hjemmet til Averys og Griffiths er at folk, som Watson og Crick, med rette har blitt hyllet for sine bidrag til molekylær genetikk, å etablere strukturen til DNA faktisk var et ganske sent bidrag til prosessen med å lære om dette spektakulært molekyl.
Strukturen til DNA
Selv om han tydeligvis ikke beskrev strukturen til DNA i sin helhet, viste Chargaff at de, i tillegg til (A + G) = (C + T), de to strengene som var kjent for å være inkludert i DNA, alltid var like fra hverandre. Dette førte til postulatet som puriner (inkludert A og G) alltid bundet til pyrimidiner (inkludert C og T) i DNA. Dette ga tredimensjonal fornuft, fordi puriner er betydelig større enn pyrimidiner, mens alle puriner i det vesentlige er i samme størrelse og alle pyrimidiner i det vesentlige av samme størrelse. Dette innebærer at to puriner bundet sammen ville ta betydelig mer plass mellom DNA-strengene enn to pyrimidiner, og også at enhver gitt purin-pyrimidin-parring ville konsumere den samme mengden plass. Å legge all denne informasjonen krevde at A binder seg til, og bare til T, og at det samme forholdet gjelder for C og G hvis denne modellen skulle vise seg vellykket. Og det har den.
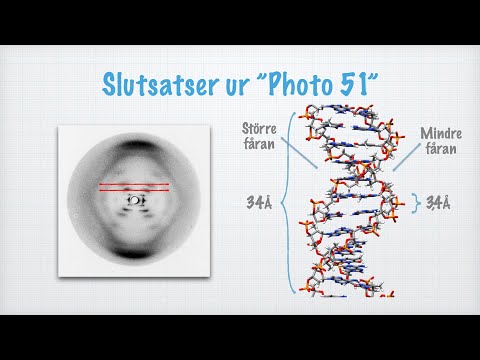
Basene (mer om disse senere) binder seg til hverandre på det indre av DNA-molekylet, som trinn i en stige. Men hva med trådene, eller "sidene", seg selv? Rosalind Franklin, i samarbeid med Watson og Crick, antok at denne "ryggraden" var laget av sukker (nærmere bestemt en pentosesukker, eller en med en 5-atom ringstruktur) og en fosfatgruppe som binder sukkerene. På grunn av den nylig avklarte ideen om baseparring, ble Franklin og de andre klar over at de to DNA-strengene i et enkelt molekyl var "komplementære", eller faktisk speilbilder av hverandre på nivået av nukleotidene deres. Dette gjorde det mulig for dem å forutsi den omtrentlige radius av den tvinnede DNA-formen i en solid grad av nøyaktighet, og røntgenstrålediffraksjonsanalyse bekreftet den spiralformede strukturen. Ideen om at heliksen var en dobbel helix var den siste store detalj om DNAs struktur som falt på plass, i 1953.
Nukleotider og nitrogenbaser
Nukleotider er de repeterende underenhetene til DNA, som er motsatsen til å si at DNA er en polymer av nukleotider. Hvert nukleotid består av et sukker kalt deoxyribose som inneholder en femkantig ringstruktur med ett oksygen og fire karbonmolekyler. Dette sukkeret er bundet til en fosfatgruppe, og to flekker langs ringen fra denne posisjonen er også bundet til en nitrogenholdig base. Fosfatgruppene kobler sukkerene sammen for å danne DNA-ryggraden, hvor de to strengene vrir seg rundt de bundne nitrogen-tunge basene midt i den doble helixen. Helixen lager en komplett 360-graders vri omtrent en gang hvert 10. basepar.
Et sukker som bare er bundet til en nitrogenholdig base kalles a nukleosid.
RNA (ribonukleinsyre) skiller seg fra DNA på tre viktige måter: Én, pyrimidin uracil er erstattet med timin. To, pentosesukkeret er ribose i stedet for deoxyribose. Og tre, RNA er nesten alltid enstrenget og kommer i flere former, diskusjonen av disse er utenfor rammen av denne artikkelen.
DNA-replikasjon
DNA blir "pakket ut" i sine to komplementære tråder når det er på tide å lage kopier. Når dette skjer, dannes datterstrenger langs ensligstående foreldre. En slik datterstreng dannes kontinuerlig ved tilsetning av enkle nukleotider, under virkning av enzymet DNA-polymerase. Denne syntesen følger ganske enkelt i retning av separasjonen av foreldre-DNA-strengene. Den andre datterstrengen former seg fra små polynukleotider kalt Okazaki fragmenter som faktisk dannes i motsatt retning av unzipping av foreldrestrenger, og blir deretter koblet sammen av enzymet DNA-ligase.
Fordi de to datterstrengene også er komplementære til hverandre, binder basene deres til slutt for å lage et dobbeltstrenget DNA-molekyl som er identisk med den overordnede.
I bakterier, som er encellede og kalles prokaryoter, sitter en enkelt kopi av bakteriene DNA (også kalt dens genom) i cytoplasmaet; ingen kjerner er til stede. I flercellede eukaryote organismer finnes DNAet i kjernen i form av kromosomer, som er sterkt oppviklede, spolede og romlige kondenserte DNA-molekyler som bare er milliondeler av en meter lange, og proteiner som kalles histoner. Ved mikroskopisk undersøkelse blir kromosomdelene som viser vekslende histon "spoler" og enkle DNA-tråder (kalt kromatin på dette organisasjonsnivået) ofte sammenlignet med perler på en streng. Noe eukaryotisk DNA finnes også i organeller av celler som heter mitokondrier.